研究活動
ヒトを含む真核生物では、転写されたRNAがプロセシングを経て成熟mRNAとなることから、転写後プロセシングの選択的な制御により、ひとつの遺伝子からでも必要に応じて多様なタンパク質が産生されている。ヒトではタンパク質遺伝子の実に9割が複数の成熟mRNAを産生することが明らかになっている。また、ヒトの疾患の原因として報告される変異のうちタンパク質の機能に影響しないものには、mRNAの転写後プロセシングに大きく影響するものが多いことが報告されている。したがって、転写後プロセシング制御機構の解明は、これまでによく研究されてきた転写調節に勝るとも劣らない重要な遺伝子発現制御機構であり、個人ゲノムの解読が進む今後の疾患研究において重要性がますます高まっていくと予想される。当研究室では、DNAから転写されたmRNA前駆体が組織特異的・発生段階依存的にプロセシングされて多様な成熟mRNAとなるための「細胞暗号」の解明を目指して研究を展開している。
- 1.蛍光選択的プロセシングレポーターによる組織特異的・発生段階依存的選択的プロセシング制御機構の解明
- 2.トランスクリプトーム解析による選択的スプライシング制御因子の標的遺伝子の網羅的探索
- 3.2つのRNA結合タンパク質による標的RNAの協働的認識機構の構造生物学的解析
- 4.リボソームタンパク質による選択的mRNAスプライシングの自己制御
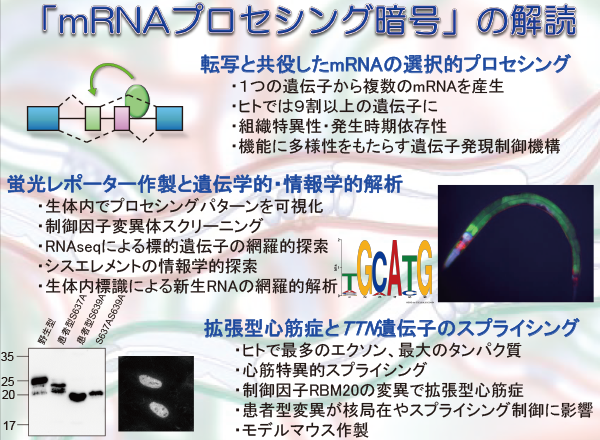
mRNAプロセシングの制御機構を生体内で解析するために、当研究室では、複数の蛍光タンパク質を用いてミニ遺伝子を構築し選択的プロセシングパターンを1細胞レベルで可視化するレポーター系を開発した(Nat Meth, 2006; Nat Protoc, 2010)。
このレポーター系を利用して、(1)線虫のFGF受容体遺伝子egl-15の筋特異的なエクソン選択性を可視化し、RBFOXファミリーRNA結合タンパク質ASD-1およびFOX-1と筋特異的RNA結合タンパク質SUP-12が協働して筋芽細胞のスプライシングを制御することでFGF受容体のリガンド特異性の制御に関わることを見出した(Nat Meth 2006; Mol Cell Biol, 2007)。(2)線虫のコラーゲン遺伝子let-2の発生段階依存的なエクソン選択性を可視化し、発生段階依存性の制御因子としてSTARファミリーRNA結合タンパク質ASD-2を同定した。さらに、選択的スプライシングによるmRNA前駆体の運命決定に重要なイントロン除去の順序を明らかにした(Genes Dev, 2008; Nat Protoc, 2010)。(3)線虫の2種類のコフィリンをコードするunc-60遺伝子の筋特異的なmRNAプロセシングパターンの切り替えをSUP-12とASD-2が協働して制御することを見出した(PLoS Genet, 2012)。(4)線虫のV-ATPaseのaサブユニットをコードするunc-32遺伝子の2組の相互排他的エクソンが組織特異的に選択されることを示し、両組の神経系特異的エクソンの選択に必須な制御因子として神経系特異的CELFファミリーRNA結合タンパク質UNC-75を同定した(PLoS Genet, 2013)。
また、線虫の相互排他的選択的エクソンについて、選択性や構造の網羅的な解析を行い、特徴を明らかにした(Worm, 2014)。
上述の研究で得られた選択的スプライシング制御因子の変異体線虫と野生型線虫のmRNAを大規模シーケンス解析して生物情報学的手法で比較することにより、選択的スプライシングパターンに差がある遺伝子を網羅的に探索し、制御因子の標的遺伝子の同定を行っている。これまでに、UNC-75の制御の標的となる合計24個の選択的スプライシング事象を同定した。さらに、スプライシングレポーター線虫の作製により、これらの標的エクソンがさまざまな組織特異的制御を受けること、UNC-75はシスエレメント(G/U)UGUUGUG配列を介して神経系特異的な制御に関わること、シスエレメントの位置により影響が異なる「位置効果」を示すことを見出した(Nucleic Acids Res, 2013)。
mRNAプロセシングを組織特異的に正確に行うためには、mRNA前駆体の塩基配列を特異的に認識して結合しプロセシングを制御するRNA結合タンパク質のはたらきが不可欠である。しかし、個々のRNA結合タンパク質のRNA結合ドメインが認識する配列は短く特異性も低いため、RNA結合タンパク質がなぜ標的遺伝子を組織特異的に正確に制御できるのか、不明な点も多い。
当研究室では、RNA結合ドメインを1つずつもつ線虫の2つのRNA結合タンパク質ASD-1とSUP-12が線維芽細胞成長因子受容体遺伝子egl-15の配列を協働的に認識する分子機構を、武蔵野大学の武藤裕教授らのグループなどとともに明らかにした(Nat Struct Mol Biol, 2014)。この成果は、分子間で特定の塩基をサンドイッチすることにより全体として安定な複合体を形成して特定の塩基配列を認識する分子機構を初めて明らかにしたもので、今後のさまざまなRNA結合タンパク質の組み合わせによる協働性の解析につながるものと期待される。
多細胞生物の選択的スプライシングでは、mRNAの途中に終止コドンを含み、品質管理機構で速やかに分解されるノンコーディングmRNAを作る例がある。この品質管理機構が欠損している線虫smg-2変異体のトランスクリプトーム解析を行って野生型株と比較し、品質管理機構の標的となる天然のノンコーディングmRNAを産生する遺伝子を探索した。その結果、リボソームタンパク質をコードする計8個の遺伝子でノンコーディングmRNAを同定した。そして、これらの遺伝子では選択的スプライシングの制御により発現量が自己制御されることを見出した。このうち60SサブユニットのL10aタンパク質をコードする遺伝子では、L10aが自身のmRNA前駆体のイントロンにあるL10ARE領域に直接かつ特異的に結合することで選択的スプライシングを自己制御すること(図参照)、L10AREの配列や位置が線虫のrpl-1遺伝子から哺乳類のRPL10A遺伝子にまでよく保存されており、哺乳類でもL10aによるL10AREを介した選択的スプライシングの自己制御があることを見出した(Nucleic Acids Res, 2016)。この研究成果は、スプライシング制御因子として機能するという、後生生物リボソームタンパク質の進化的に保存された新たなリボソーム外での機能を明らかにしたものである。
難病基盤・応用研究プロジェクト室「難病筋疾患」タイチン遺伝子のスプライシング異常と拡張型心筋症
拡張型心筋症は、心筋壁が薄く伸展することによって心室の内腔が拡大しポンプ機能が障害されて機能不全に陥るものであり、根本的な治療法が確立されていない。近年、拡張型心筋症患者の遺伝子解析により、心筋サルコメアを構成し筋収縮に関連するさまざまなタンパク質の遺伝子変異が相次いで報告されている。タイチンもその1つであり、サルコメアが伸展した際に受動張力を発揮して過伸展を防ぐ。心臓では、タイチンをコードするTTN遺伝子の選択的スプライシングにより短いN2B型とやや長いN2BA型のタイチンが発現するが、拡張型心筋症心筋では、張力が大きいN2B型の比率が減少し張力が小さいN2BA型の比率が増加する。拡張型心筋症患者の約25%でTTN遺伝子の短縮変異やミスセンス変異が見つかるが、これらの変異の機能的意義は必ずしも明らかではない。一方、RNA結合タンパクRBM20の欠損ラットが心筋でTtn遺伝子のスプライシング異常を示して心筋症を自然発症することが近年報告された。
当プロジェクトでは、TTN遺伝子の二色蛍光スプライシングレポーターミニ遺伝子を作製し、RBM20によるTTN遺伝子の心筋型スプライシング制御を解析する実験系を構築した。そして、拡張型心筋症患者で変異が集中して報告されているRBM20のRSRSP配列中のSer635残基とSer637残基がともにリン酸化されること、それがRBM20の核移行に必須であることを見出した。
当プロジェクトのメンバーである分子病態分野の木村彰方教授らは、木村らは遺伝性心疾患関連67遺伝子の変異をスクリーニングするシステムを構築し、家族歴を有する拡張型心筋症患者に新たにRBM20のR634W変異を見出している。また、欧米人患者では、D888N、G1031X、P1081R、E1206Kなどのミスセンス変異やナンセンス変異が報告されている。そこで、上述のレポーターミニ遺伝子を利用して、これらの変異がRBM20のスプライシング制御能に与える影響を順次解析中である。また、タイチンのN2B型とN2BA型の発現比率の変化と心筋症の因果関係の実験的に検証するための遺伝子改変マウスや、Rbm20遺伝子に患者型のアミノ酸置換変異を導入するモデルマウスの作製を並行して進めている。当プロジェクトにより拡張型心筋症とTTN遺伝子のスプライシング制御機構の関連が明らかになることで、拡張型心筋症の病態解明や治療標的の開発につながることが期待される。
教育活動
黒柳 秀人:大学院医歯学総合研究科、医学部保健衛生学科、東京大学非常勤講師(教養学部後期課程)
和仁翔太郎:歯学部歯学科